中国农大新闻网讯 近日,中国农大农学院李自超和张洪亮团队在著名期刊《基因组生物学》(Genome Biology)上发表了题为“大规模水稻杂种基因组及转录组揭示杂种优势的核心机制”(Large-scale genomic and transcriptomic profiles of rice hybrids reveal a core mechanism underlying heterosis)的研究论文。该研究通过大规模的杂种群体表型、QTL、转录组、动力学模拟以及酵母实验验证等技术手段,系统解析了水稻杂种优势的遗传基础,并提出了杂种优势发生的核心机制-不充分背景下纯合子功能不足模型(HoIIB)。该模型能够解释大多数已知的杂种优势假说和现象,为水稻遗传改良和杂交育种提供了新的理论支持。我院毕业博士谢建引、湖南杂交水稻研究中心的王伟平研究员和我院博士生杨涛为论文的第一作者。我院张洪亮教授、李自超教授为论文的通讯作者。生物学院张子丁教授和农学院王向峰教授等对研究工作进行了指导和帮助。
杂种优势是生物界普遍存在的现象,已在农业生产中得到了广泛应用。一个多世纪以来,杂种优势机理一直是生命科学领域的热点问题。科学家很早就提出了显性、超显性和上位性假说来解释杂种优势形成的遗传机理,现代的酶学理论及以其为基础发展而来的代谢组模型则从生理生化反应的角度对杂种优势的产生机理进行了阐述。最近几十年,随着分子生物学和测序技术的快速发展,大量研究人员以拟南芥、水稻、玉米和油菜等作物为研究对象,对杂种优势的成因在不同层面开展了详细研究,极大地丰富了我们对农作物杂种优势的认识。然而,已有研究更多解释的是杂种优势现象的表现形式,人们尚未明确杂种优势形成的内在机制,包括很多学说或模型成立的前提--非加性遗传效应形成的内在机制也不十分清楚。
1.杂种优势以及非加性效应的形成高度依赖环境以及遗传背景
该研究中,作者通过对418个水稻杂种组合及其亲本,2808份玉米杂种组合及其亲本,及1600余份小麦杂种组合及其亲本在多个环境下的表型分析,发现不同环境对杂种的影响普遍强于自交系亲本,不同组合类型、不同性状以及不同环境杂种优势表现差异明显,表明杂种优势的表现高度依赖于环境。分析全基因组关联分析检测到的亲本、杂种及中亲优势相关QTL及其遗传效应(加性、显性、超显性),发现非加性QTL相比于加性QTL对不同环境或遗传背景的变化更为敏感。对优良杂交组合两优培九及其双亲(培矮64S和9311)的幼穗转录组数据分析表明,非加性表达条件下对遗传背景(其转录因子)的依赖性高于加性表达条件下(图1)。上述结果表明,杂种优势以及非加性效应的形成高度依赖环境以及遗传背景。

图1. 转录水平非加性与加性现象对背景依赖性比较
2.提出了杂种优势形成的核心机制-HoIIB模型
为明确遗传背景与杂种优势及非加性效应的内在关系,作者利用希尔方程系统模拟了不同场景中不同配体浓度下受体在双亲及其杂种中的反应,模拟结果进一步从理论上说明非加性及杂种优势的形成与遗传背景密不可分。基于以上结果,作者提出不充分背景下的纯合子功能不足是非加性及杂种优势形成的核心机制,并将其命名为HoIIB(Homo-insufficiency under insufficient background)模型(图2)。
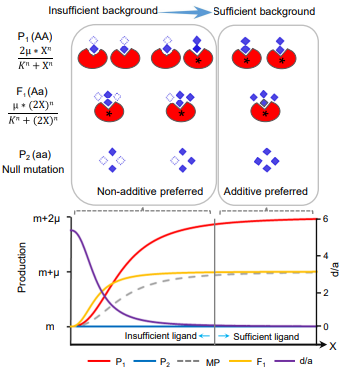
图2 存在/缺失等位变异在不同背景供应条件下的杂种优势表现
该模型认为,非加性效应不是一个基因的固有特征;其本质是亲本中两个纯合等位基因在背景不充分的情况下功能未能发挥,而在同样背景下杂种F1中某一个等位基因可以充分发挥,从而表现出部分显性、显性甚至超显性。理论上,HoIIB模型应有三个基本特征:(1)非加性(包括显性和超显性)条件下,强功能的纯合子会表现出功能发挥不足;(2)同一位点两个等位基因之间的累加或互补更容易形成超显性;(3)非加性状态下,目标基因的功能对背景的响应更强。三个特征在大规模杂交组合表型、QTL以及转录组水平均得到了印证(图3a-d)。
根据HoIIB模型,既然非加性的形成与遗传背景有关,理论上非加性与加性之间可以因遗传背景的不同而发生转换。作者在二倍体酵母体系实验中验证了该推断。实验结果显示,在野生型中目标基因SSU1在转录水平及其对酵母生长的效应方面均表现出超显性,当上调其背景(即其转录因子)FZF1的表达时,随着FZF1转录水平增强,目标基因SSU1的显性度逐渐降低而接近加性(图3e-f)。对水稻产量相关基因Hd3a的分析同样印证了该模型。
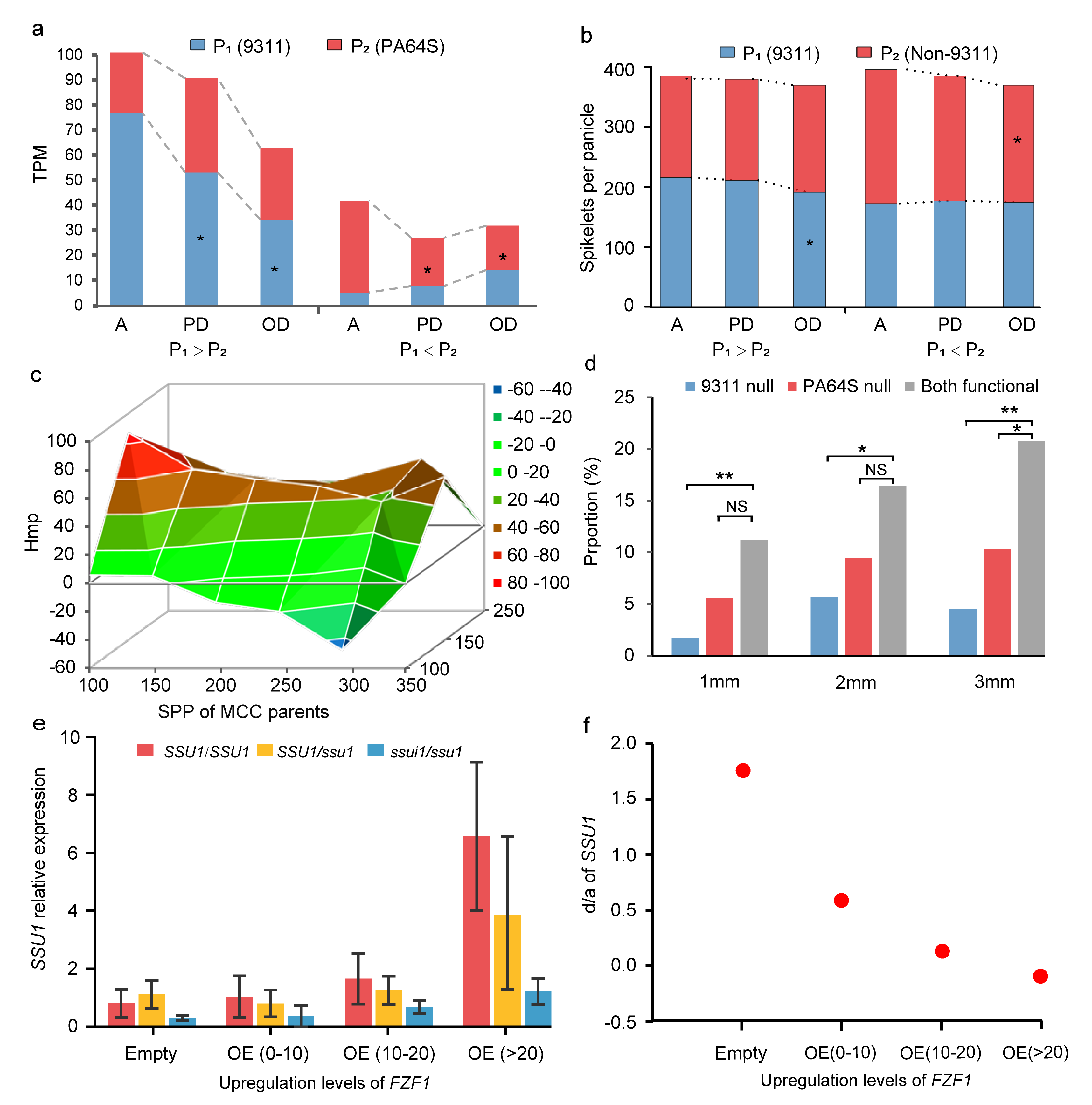
图3 转录、QTL和表型水平上背景效应对加性、显性和超显性现象的影响
3.HoIIB模型对杂交育种的理论意义
讨论中作者探讨了HoIIB模型对未来水稻遗传改良和杂交育种的理论意义。首先,该研究表明,显性或杂种优势不是目标基因本身的内在特性,而是其背景赋予的特性。因此,当一个基因或组合表现出明显的杂种优势现象时,表明该表达调控体系中存在明显短板;此时,利用杂种优势只是这个调控体系的次优解。找到调控体系的短板,加以改良从而释放纯合子的最大功能才是育种的终极目标。其次,当一个基因或因子在不同背景或条件下表现出稳定的加性效应时,该基因或因子很可能是一个系统性短板。该研究为寻找某个物种或群体的系统性短板,从而改良该系统性短板培育突破性品种提供了理论依据。第三,该理论的提出并非意味着不再利用杂种优势。相反,作者分析认为杂种优势的利用在相当长的时间内仍将是重要的育种策略。最后,该理论还有助于理解一般配合力和特殊配合力及其在育种中的利用,并提出了一个有效培育突破性杂交品种的策略:持续聚合系统性限制因子的优异等位基因,并通过传统和生物技术手段将其改良,以持续改良基因组背景;将更多背景效应引入杂交育种的基因组选择模型,从而改进对特殊配合力预测的准确性。
论文链接:https://genomebiology.biomedcentral.com/articles/10.1186/s13059-022-02822-8
供稿:农学院
供图:农学院
编辑:马文哲
责编:于哲


